Rife machines, Cancer Viruses SV-40, a pro-cancer virus in vaccines
In 1955, Jonas Salk performed a medical miracle when he discovered how
to mass
produce polio vaccine by growing it on the kidneys of rhesus monkeys.
While there
is no question that thousands were saved from the ravages of polio by
the Salk vaccine,
by 1960 a problem had surfaced -- researchers had isolated a viral contaminate
in the
vaccine, Simian (monkey) Virus # 40. It seems that when the live polio
virus grown on
monkey tissues was extracted for vaccine production this SV-40 virus was
extracted
as well.
When SV-40 was injected into research animals it produced brain cancer.
It appears
our government didn't wish to create a public panic or discredit the public
health
service, because instead of recalling the tainted vaccines, it quietly
ordered the
manufacturers to find a monkey free of SV-40 and continue production.
As of 1963
, the rhesus monkey had been replaced with the African green monkey for
production
of a safer polio vaccine, but between the years of 1955 and 1963 as many
as 98 million
Americans had received doses of live polio virus vaccines tainted with
SV-40.
Nowadays SV-40 has appeared in 61% of all new cancer patients -- patients
too young
to have received the contaminated vaccine being administered forty years
ago who are
now believed to have been infected by human to human transmission. Being
a blood
born organism, it is also suspected that SV-40 is transmissible from mother
to child
during pregnancy. The more this matter is researched the more startling
the evidence.
Senior epidemiologist at the National Institutes of Health, Dr. Howard
Strickler,
has plotted a geographic pattern to the cancers associated with SV-40
helping to confirm
its link to the tainted vaccine. People who lived in Massachusetts and
Illinois who
received identified lot numbers of the contaminated vaccine administered
in the 1950s
are now demonstrating ten times the rate of the osteosarcoma bone tumors
as those
who received vaccine free of the SV-40 contaminate in other parts of the
country.
DNA Polyoma Viruses
In 1964, studies were conducted on a polyoma virus (a tumor-producing
DNA virus).
It was discovered that the persistent genetic DNA material in the polyoma
virus
brought about malignant transformations in hamster embryo cell cultures.
This was
reported in the November 23, 1964 issue of the Journal of the American
Medical
Association.
SV-40 is one example of a DNA polyoma virus. Polyoma (many tumor-causing)
viruses cause prolonged infection where tissue is destroyed, integrate
into the
hosts genetic material, are capable of mutating a cell, may reproduce
after coming
into contact with a 'helper' virus, enable the separate replication of
the viral
genome, can generate immune responses, and they can induce malignancy.
Scientists are amazed at how little genetic information these viruses
carry in
proportion to the damage they can cause.
The 'D' in DNA and the 'R' in RNA have characteristics which are dependent
on
the kind of sugar molecule associated with it. DNA exists predominantly
in the
nucleus, but is also represented in the cytoplasm and in the mitochondria.
RNA
is also present in the cytoplasm. When viral RNA or DNA combines with
the
genetic material in the cell itself, the viral genetic material can become
part of
the host cell genetic code, altering the genetic structure of the cell.
When the
altered cell duplicates, the encoded viral genetic material may affect
cellular
processes in such a way as to produce abnormal cells, which sometimes
become
malignant or cancerous.
Cancer-Causing RNA Viruses and DNA proviruses
The discovery in 1975 that viruses causing cancer in animals had a special
enzyme
called reverse transcriptase makes the problem even more interesting.
These kind
of viruses are called RNA viruses. When an RNA virus has the reverse transcriptase
enzyme within its structure, it allows the virus to actually form strands
of DNA
which easily integrate with the DNA of the host cell which it infects.
Studies by
Dr. Robert Simpson of Rutgers University indicate that RNA viruses which
do
not cause cancer can also form DNA, even without the presence of reverse
transcriptase. DNA formed in this way from an RNA virus is called a provirus.
It is known that some non-cancerous viruses have a tendency to exist as
proviruses
for long periods of time in cells without causing any apparent disease.
In other words,
they remain latent. Some examples of common RNA viruses that do not cause
cancer, per se, but have the capacity to form proviruses are influenza,
measles,
mumps and polio viruses.
Viruses as Catalysts for Cancer
An article in the January 6, 1962 Science Newsletter indicated that 'common
human
viruses act as carriers in causing cancer by interacting with cancer-causing
chemicals;
this has been indicated by experiments which show that cancer-causing
substances
that are present in too small a quantity by itself will become active
and create tumors
when combined with single doses of virus. Malignant tumors appeared in
five type
of injected mice.' The viruses mentioned were ECHO9, B-4, Coxsackie, and
Polio
virus 2. The article further indicated that 'viruses may also activate
other cancer
causing substances besides chemicals in the environment, such as DMBA,
AF,
and DBA.'
Even common non-tumor viruses, including those in smallpox vaccine and
polio virus
2, can act as carcinogens. It was reported in Science on December 15,
1961 that
these common viruses acted as catalysts in producing cancer when given
to mice
in combination with known organic carcinogens in amounts too small to
induce
tumors themselves. This means that some vaccinations will induce cancer,
when
combined with the growing problem of environmental pollution from toxic
by-products
of agriculture (pesticides on and in food) and industry.
A Listing of Cancer Causing Microbes
The July 14th 1997 issue of Business Week has an article in it about
how many
cancers are being linked to various viruses, and bacteria ,and parasites.
Among the organisms now linked to cancer are as follows
Microbe Type of Cancer
------- --------------
Hepatitis B Virus Liver Cancer
Human Papiloma Virus ( HPV ) Cervical Cancer
Helicobacter Pylori Stomach Cancer
HTLV-1 A type of Leukemia in Japan
Epstein- Barr virus (EBV) Burketts Lymphoma, naso pharyngeal
cancer
Kaposi's Sarcoma Herpes Virus (KSHV) Kaposi's Sarcoma, and 100 % of
Myeloma cases.
Schistosomiasis Bladder Cancer
Liver Flukes Liver and biliary cancer
Helicobacter hepaticus Liver cancer
Hepatitis C Virus Liver cancer
Papillomaviruses (HPV-5,HPV-8,HPV-17) Skin cancer
Polyomavirus (BK and JC) Neural tumors? and insulinomas?
Retrovirus (HTLV-2) Hairy-cell leukemia
Lyme Disease bacteria B. Burgdorferi Skin and Breast cancer
Epstein-Barr Virus Majority of Non-Hogkins lymphoma (sp)
Granuloma type Virus Skin Cancer (Not confirmed)
References
Fisher, B. L. (1997). Workshop on Simian Virus 40: A Possible Human
Polyomavirus. National Vaccine Information Center, January 27, On-line
at http://www.909shot.com/polio197.html
Carbone, M., et al. (1996). SV-40 Like Sequences in Human Bone Tumors.
Oncogene, 13(3), 527-535.
Elswood, B. F., & Stricker, R. B. (1995). Polio Vaccines and the
Origin of AIDS.
Medical Hypotheses, 42(6), 347-354.
Krieg, P., Amtmann E, Jonas, D., Fischer, H., Zang, K., & Sauer G.
(1981).
Episomal Simian Virus 40 Genomes in Human Brain Tumors. Proceedings
of the National Academy of Sciences of the United States of America, 78(10),
6446-6450.
Lednicky, J. A., Garcea, R. L., Bergsagel, D. J., & Butel, J. S.
(1995). Natural
Simian Virus 40 Strains are Present in Human choroid Plexus and Ependymoma
tumors. Virology, 212(2), 710-717.
Martini, F., et al. (1995). Human Brain Tumors and Simian Virus 40. Journal
of
the National Cancer Institute, 87(17), 1331.
Martini, F., et al. (1996). SV-40 Early Region and Large T Antigen in
Human
Brain Tumors, Peripheral Blood Cells, and Sperm Fluids From Healthy Individuals.
Cancer Research, 56(20), 4820-4825.
Pass, H. I., Kennedy, R. C., & Carbone, M. (1996). Evidence for and
Implications
of SV-40 Like Sequences in Human Mesotheliomas. Important Advances in
Oncology, 89-108.
Rock, A. (1996). The Lethal Dangers of the Billion Dollar Vaccine Business.
Money, December, pages 148-163.
Tognon, M., et al. (1996). Large T Antigen Coding Sequences of Two DNA
Tumor Viruses, BK and SV-40, and Nonrandom Chromosome Changes in
Two Glioblastoma Cell Lines. Cancer Genetics and Cytogenics, 90(1), 17-23.
Inherited virus may play role in breast cancer
NEW YORK, Aug 12 (Reuters Health) -- An inherited virus may be one of
the
factors that triggers breast cancer in humans, researchers report. Scientists
say that a primitive retrovirus, human mammary tumor virus (HMTV), has
been
identified in human breast cancer tissues.
'If a definitive link to this retrovirus is established, HMTV may become
a target
for a vaccine to prevent breast cancer and a target for new treatments
for breast
cancer,' explained study lead author Dr. Robert Garry of Tulane University
in New
Orleans, Louisiana.
A retrovirus similar to HMTV has already been linked to malignant breast
tumors
in mice. Speaking to attendees at the 11th International Congress of Virology
in
Sydney, Australia, Garry explained that vertebrate species other than
mice --
including some humans -- carry similar viruses.
He said that his team had identified the virus, dubbed HMTV, in breast
cancer
tissue and other organ issue from breast cancer patients, and also in
tissues from
individuals who did not have breast cancer.
The virus, he said, is likely to be 'a cofactor' for triggering breast
cancer, along
with other factors such as an individual's genetic makeup. Dr. Orli Etingin,
an
oncologist and assistant professor of medicine at New York Hospital/Cornell
Medical Center in New York City, called the finding 'a very interesting
new
piece of the (cancer) puzzle.'
Speaking with Reuters Health, she noted that 'retroviruses have (already)
been implicated in certain kinds of lymphomas.' But she believes that
'a lot more
research really has to be done in order to confirm the finding and also
to establish
what the relationship of the virus is to the development of tumors in
humans.'
Subcellular Life Forms
John Baez
September 3, 1998
Also available in Postscript and LaTeX, thanks to Stephen Mulraney.
I like biology, but as a mathematician, I am drawn to the elegance of
the very
simplest forms of life: the subcellular life forms. They are so simple,
in fact,
that even calling them "alive" can be controversial. They lack
many of the
usual features of life. They don't have cell walls, most of them don't
metabolize,
and they are all parasitic, depending on other organisms for their ability
to reproduce!
Some of them even have no genetic code! Many of them cause diseases, but
others
are crucial to the well-being of their host, and many are so well integrated
with their
host that it becomes difficult to decide whether they are part of the
host or a
separate entity.
Indeed, besides my love of elegance and my morbid fascination with parasites,
the main reason subcellular life forms appeal to me is that they challenge
our
naive notion of organisms as entities with clear, well-defined boundaries.
It's
clear by now that life doesn't respect this simple picture. Whenever a
pattern
of any sort, however abstract, is able to replicate itself, it does! Typically
these
patterns overlap and interact in subtle ways, so one can't easily say
where one
ends and the other begins.
These are the main kinds of subcellular life forms that I know about so far:
Viruses
Viroids
Virusoids
Plasmids
Transposons
and
Prions
I'll say a little about each kind. If you know any more fascinating facts
about
subcellular life forms - especially if you know kinds that aren't on this
list -
please email me!
Some of the above terms are defined in an essay by Diener and Prusiner
called
"The Recognition of Subviral Pathogens" [MM]. But beware! People
argue quite
a bit about the correct classification of these life forms. That's part
of what's
interesting about them: they really stretch our ideas in biology to the
breaking point.
One thing to keep in mind: these life forms are small. Remember that
DNA is
a double helix containing information in the form of AT and CG "base
pairs"
(paired molecules of adenosine and thymine, or cytosine and guanine),
while
single-stranded RNA is a single helix containing information in the form
of
A, U, C, and G "bases" (molecules of adenosine, uracil, cytosine
and guanine).
The human genome is made of DNA and contains about 5 billion base pairs.
The genome of a bacterium is also made of DNA but has less than 10 million
bases. The potato spindle tuber viroid, on the other hand, is nothing
but a
circular loop of RNA consisting of 359 bases! Small, simple - but effective!
--------------------------------------------------------------------------------
Viruses
Diener and Prusiner define a virus to be a "small infectious pathogen
composed
of one or more nucleic acid molecules usually surrounded by a protein
coat."
They typically reproduce by latching onto the wall of a cell and inserting
their
genetic material - i.e., the nucleic acids - into the cell. This genetic
material
then uses the cell's machinery to make more copies of the virus. Typically,
these copies overrun the cell until it bursts. However, the actual life
cycle
of a virus is often more complicated than this thumbnail sketch! Viruses
use
a large number of sneaky tricks to overcome the defense mechanisms of
the cell.
Apart from their intrinsic interest, viruses are important because they
cause
many diseases among humans, such as:
the common cold
influenza (the flu)
measles
rubella
mumps
warts
chickenpox
smallpox
acquired immunodeficiency syndrome (AIDS)
herpes
hepatitis
rabies
poliomyelitis (polio)
encephalomyelitis
encephalitis
yellow fever
dengue fever
as well as diseases of domesticated animals and plants.
There is standard taxonomy of viruses [CT] [Ma2], [F], but I will content
myself with a rough classification into the following 3 sorts:
DNA viruses
RNA viruses
and
Retroviruses
DNA viruses
The genome of a DNA virus is a single molecule of DNA, either linear or
circular,
and usually double-stranded. Outside the host cell, this DNA is surrounded
by a
protein coat. There are 5 known families of DNA viruses affecting humans.
The size and structure of the DNA viruses varies widely, from the small
hepatitus
B virus (HBV), whose round shell contains a circular DNA molecule with
about
2,400 base pairs, to the large brick-shaped or ovoid pox viruses, which
have a
lipid coating and whose DNA has between 120,000 and 360,000 base pairs.
Like retroviruses, some DNA viruses work their way into the nucleus of
their
host cell and then copy themselves into the host's DNA. An example is
the
hepatitus B virus, which occupies liver cells. This can cause tumors.
RNA viruses
The genome of an RNA virus is usually a single molecule of RNA, either
linear or
circular, but some contain up to a dozen molecules of RNA. Outside the
host cell,
this RNA is protected by a protein coat. Most viruses are RNA viruses.
There
are 13 known families of RNA viruses affecting humans. RNA virus range
widely
in morphology and size, with their genome containing anywhere from 1,700
to 60,000 nucleotides.
(Actually the smallest one, the hepatitis delta agent (HDV), is quite
different
from all the rest. Like a virusoid, it is a circular loop of RNA that
can only
reproduce in cells infected by a helper virus, the hepatitus B virus.
But unlike
a virusoid, it affects animals rather than plants, it has its own protein
coat,
and its genome is bigger than that of a virusoid, having 1,700 nucleotides
instead of a mere 350 or so. However, its genome is much smaller than
that
of any other virus.)
One can broadly classify RNA viruses into:
positive-strand RNA viruses
negative-strand RNA viruses
double-stranded RNA viruses
A "positive-strand" RNA virus consists of single-stranded RNA
that functions
directly as messenger RNA in the host cell, so that ribosomes in the host
cell
synthesize various proteins needed by the virus when encountering this
RNA.
A "negative-strand" RNA virus consists of single-stranded RNA
that does
not function as messenger RNA, since it contains the complementary base
pairs.
Negative-strand RNA viruses carry enzymes with them into the host cell
to
synthesize messenger RNA from the RNA in the virus. "Double-stranded"
RNA viruses have both positive and negative strands. For some reason these
are more likely to consist of several separate pieces of RNA.
Retroviruses
Retroviruses are like RNA viruses when outside the host cell, but once
inside the cell's nucleus, they can copy themselves into the DNA of the
host
cell using an enzyme called "reverse transcriptase", which translates
RNA
into DNA. They are thus intermediate between RNA viruses and nuclear DNA
viruses. Once they are integrated into the DNA of the host cell, they
may
take a long time to reemerge. In fact, so-called "endogenous retroviruses"
can be passed down from generation to generation, indistinguishable from
any
other cellular gene, and evolving with their hosts! The very distinction
between
host and parasite becomes somewhat blurry in this case. In fact, once
an
endogenous retrovirus lost the genes that coat for its protein coat, it
would
become indistinguishable from an LTR retrotransposon - one of the many
kinds of "junk DNA" cluttering up our chromosomes.
It has been estimated that between .01% and .1% of the genome of wild
and
laboratory mice consists of endogenous retroviruses. The same is probably
true for humans. to form protein coats - since most mammalian DNA serves
no known purpose, the above figures may be drastic underestimates. Indeed,
97% of human DNA is so-called "junk" DNA of this sort!
Retroviruses are important in genetic engineering because they raised
for the
first time the possibility that RNA could be transcribed into DNA, rather
than
the reverse. In fact, some of them are currently being deliberately used
by
scientists to add new genes to mammalian cells.
In addition, retroviruses are important because AIDS is caused by a retrovirus:
the human immunodeficiency virus (HIV). This is part of why AIDS is so
difficult
to treat. Most usual ways of killing viruses have no effect on retroviruses
when
they are latent in the DNA of the host cell.
Many retroviruses cause tumors in animals. These viruses contain host-derived
genetic information.
--------------------------------------------------------------------------------
Viroids
A viroid is defined to be a "small infectious pathogen composed entirely
of a low
molecular weight RNA molecule". Thus, unlike a virus, a viroid has
no protein
coat. It is nothing but a single-stranded circular loop of RNA! Most viroids
consist
of about 250 to 575 nucleotides, much smaller than a typical virus. Also,
viroids don't
function as messenger RNAs, so they don't make the cell synthesize enzymes:
they rely completely on pre-existing enzymes in the host for their reproduction.
Most known viroids cause diseases in plants. The first viroid was discovered
in 1971,
by Diener. It's called the potato spindle tuber virus (PSTV), since it
causes a disease
that makes potatos abnormally long and sometimes cracked. At the time,
Diener's
isolation of the viroid causing this disease met with some skepticism,
since it was
so much smaller than any known virus. By 1991, however, at least 15 plant
diseases
had been traced to viroids. There are also 2 viroids known, the hop latent
viroid (HLV)
and a viroid living in grapevines, that cause no known symptoms! This
raises the
fascinating possibility that there could be more such viroids lurking
around.
The complete molecular structure of many viroids has been worked out,
which
has allowed a classification of viroids on the basis of their RNA sequences.
Roughly speaking, there are a large family of viroids that share many
features
with PSTV, together with one viroid that seems very different: the avocado
sunblotch viroid (ASBV). McInnes and Simons have proposed a further classification
of the PSTV-type viroids into three kinds [Ma1].
It is clear from these RNA sequences that viroids are not "degenerate
viruses",
as had once been thought. They are quite different from any known viruses.
One interesting theory is that they arose from RNA that escaped from cell
nuclei.
It's also interesting that all viroid diseases have been detected in
the 20th century,
some quite recently - in contrast to diseases caused by viruses. Also,
many viroid
diseases have been spreading after their discovery, often due to human
activity.
A fascinating example is the coconut cadang-cadang viroid (CCCV), a disease
of coconuts which has been spreading throughout the Phillipines. On the
island
of Luzon, a puzzling feature of this disease was that it only affected
crops owned
by speakers of Bicalano, while adjacent crops owned by speakers of Tagalog
went unharmed! Eventually people realized that the viroids were spread
by workers
cutting the palms. Tagalog owners prefer to hire Tagalog workers, while
Bicalanos
hire Bicalanos, some of whom came from an area where the disease was
prevalent. (See the article by Maramarosch entitled "The Cadang-Cadang
Viroid Disease of Palms" [D].)
Because of this sort of epidemiology, Diener has suggested that viroids
may
be latent to their native host plants (like HLV), becoming pathogenic
only
when transferred to other species thanks to agriculture. Indeed, the viroid
causing tomato "planta macho" disease in Mexico, TPMV, has also
been
found in wild plants there, where it seems sometimes "recover"
from ASBV
by sending up a new shoot. This new shoot is still infected with the viroid,
but
it shows no symptoms other than reduced fruit yield. Descendents of such
a
"recovered" tree are also infected with the viroid, and also
symptomless,
except for reduced fruit yield. Thus the avocado appears able to "come
to
terms" with the viroid in some way. Personally, I'd like to raise
this possibility:
that some viroids actually play a beneficial role in their native host
plants!
This may seem surprising, but when we compare the behavior of plasmids,
it may seem less so.
--------------------------------------------------------------------------------
Virusoids
A virusoid is a "viroid-like RNA encapsidated in a virus shell that
also contains viral
RNA". In other words, like viroids, they are circular loops of RNA,
usually containing
about 350 nucleotides. But unlike viroids, they reside inside the protein
coat of a
"helper virus". They can only reproduce in cells that have been
infected by this
helper virus, because they use some of the RNA of the helper virus to
reproduce.
The helper virus is typically an RNA virus consisting of about 4500 nucleotides.
In short, a virusoid is a parasite of its helper virus. But it's not
always so simple.
Sometimes the helper virus is unable to reproduce unless the virusoid
is present!
Then we have symbiosis rather than parasitism.
The first virusoids were discovered in the early 1980s in Australia,
associated with
viruses causing plant diseases such as velevet tobacco mottle (VTMoV),
solanum
nodiflorum mottle (SNMV), lucerne transient streak (LTSV), and subterranean
clover mottle (SCMoV).
An interesting theory about the origin of virsoids is that in plants
infected with both
viruses and viroids, the viroids got encapsidated in the viruses and later
lost their
ability to reproduce independently.
At this point, I should admit that the terminology concerning virusoids
is quite confusing
to me. People sometime use "satellite RNA" as a synonym for
"virusoid", but I'm not
always sure when it's supposed to be an exact synonym. Diener and Prusiner
define a
"satellite RNA" to be a "small RNA that does becomes packaged
in protein shell made
from coat proteins of another, unrelated, helper virus, on which the satellite
RNA
depends for its reproduction". The similar-sounding term "satellite
virus" appears
to be reserved for an RNA virus that depends for its reproduction on an
unrelated
helper virus, but whose genome codes for its own protein coat.
--------------------------------------------------------------------------------
Plasmids
A plasmid is a "small autonomously replicating circular molecule
of DNA that is
devoid of protein and not essential for the survival of its host".
Plasmids range in
size greatly, from about 4350 to 240,000 base pairs. Most known plasmids
infect
bacteria, but some infect plant and animal cells. They often copy themselves
into
the DNA of the host cell, and many carry genetic traits from one cell
to another.
Most plasmids keep a limit on the number of copies of themselves they
keep in
each host - the so-called "copy number", which ranges from 1
to about 40. Many
plasmids are "conjugative". This means they can transfer copies
of themselves
from one host to another by forcing the host to undergo "conjugation"
- a form of
sex in which genetic material is exchanged between bacteria.
People tend not to speak of plasmids as "life forms" quite as
often as they do with
viruses. In part this may be because plasmids are sometimes beneficial
to their host
cells, rather than pathogenic.
However, is difficult for me to resist the impression that plasmids are
just as
" alive" as viruses. Indeed, some viruses become plasmids when
parts of them
are missing! For example, the "lambda bacteriophage" is a virus
that infects the
intestinal bacterium E. coli, but "lambda dv particles", which
arise from the lambda
phage simply by deleting some DNA, are plasmids. The lambda phage multiplies
inside its host and then kills it by "lysis", which destroys
the cell membrane and
releases lots of copies. The lambda dv particles, on the other hand, stays
in the
cell in a fairly stable number of copies and does not kill its host. The
difference
is that while the lambda dv particles contain genes for replication, they
lack genes
for lysis and the protein coat.
If we think of plasmids as life forms, we must admit that they are very
successful.
Many plasmids spread so thoroughly in cultures of bacteria that less than
one cell
in 100,000 lacks a copy! Some kinds of plasmids contain genes that help
make
sure copies are efficiently passed on to both daughter cells when the
host cell divides.
F plasmids have a particularly clever mechanism - they temporarily inhibit
cell division
when they have not yet replicated inside the host!
Plasmids are diverse and very interesting. Some important kinds are:
R Plasmids
F Plasmids
Colicin Plasmids
Virulence Plasmids
Metabolic Plasmids
Tumor-Causing Plasmids
and
Cryptic Plasmids
While they don't quite fit under this heading, I can't resist also mentioning
Cosmids
and
Phasmids
These are man-made entities based on plasmids, used in biotechnology.
Are they alive?
You judge.
Some good books on plasmids include "Plasmids" by Paul Broda
[B], "Bacterial Plasmids"
by Kimber Hardy [H], and "Plasmids of Eukaryotes: Fundamentals and
Applications"
by K. Esser et al [E].
R Plasmids
R plasmids were first discovered in Japan in 1957. In Japan, dysentary
was treated
with sulphonamide until about 1950. Then, more and more strains of the
bacteria
causing dysentary became resistant to this antibiotic, rapidly rendering
it ineffective.
Doctors then began using tetracycline, streptomycin and chloramphenicol.
By 1957,
2% of the bacteria causing dysentary were resistant to one of more of
these drugs,
and by 1960, 13% were resistant. It turned out that R plasmids were the
culprit!
R plasmids contain genes that give their bacterial hosts resistance to
antibiotics as
well as to poisonous metal ions such as arsenic, silver, copper, mercury,
lead, zinc
and so on. Because many R plasmids are conjugative, this resistance can
spread
from one bacterium to another. Because they can live in more than one
species of
bacteria, R plasmids can also spread resistance between bacteria of different
species!
Spread of resistance to antibiotics is now a major problem in medicine.
Drugs which
were used for many years to control bacterial diseases are now becoming
helpless
against new resistant strains. The problem has been made worse by the
tendency
for doctors and veterinarians to use antibiotics when they aren't strictly
necessary,
for example as part of livestock food. As a result an environment is created
where
bacteria with resistance have a great competitive advantage, so they spread
rapidly.
It has also recently been found that weeds growing near crops that were
genetically
engineered to resist herbicides can acquire this trait. I'm not sure,
but I suspect that
this happens via plasmids as well.
R plasmids make it clear that the idea of evolution as a battle between
species with
separately evolving genomes is a great oversimplification. Instead, genetic
communication and cooperation between different species can be very important.
F Plasmids
F plasmids live in the bacterium E. coli and were discovered in the 1920s.
An F plasmid
contains genes that make the cell membrane of its host form long tubes.
These tubes,
called "sex pili", attach themselves to other E. coli and puncture
their cell membranes.
The F plasmid then duplicates and a copy passes from the original host
to the new host.
A clever system has evolved to ensure that the sex pili of a given bacterium
never attach
to itself.
F plasmids give their hosts no known traits besides these sex pili. The
evolutionary origins
of sex are much debated these days; we see here the fascinating possibility
that sex can
originate as a kind of disease whose sole function is to spread a parasite!
Colicin Plasmids
Colicin plasmids contain genes that give their host bacterium a certain
small probability
of bursting open and releasing chemicals called "colicins".
These chemicals kill other
bacteria by rendering their cell membranes permeable to important ions.
There are many
strains of colicin plasmid. Each one confers immunity only to the particular
sort of colicin
it produces. Different strains of colicin plasmid are "incompatible",
meaning that a given
strain bacterium cannot stably contain both.
In short, different strains of colicin plasmid compete with each other
using the resources
of their hosts. A colicin plasmid will confer an advantage to its host
bacteria if the other
strains of bacteria nearby do not have a colicin plasmid. However, when
there are many
different strains of colicin plasmid present, all strains of host bacteria
suffer. Thus there
is a certain similarity between colicin plasmids and "protection
rackets" run by Mafia-like
gangs.
Colicin plasmids are not the only sort of plasmids that exhibit incompatibility.
Similar
plasmids tend to be incompatible with each other, while drastically different
plasmids
are usually compatible. One theory is that incompatible plasmids use the
same mechanisms
to maintain their copy number. In a cell containing two incompatible sorts
of plasmid, their reproduction is blocked until the total number of copies
of the two together drops to the
copy number of each one. This is an unstable situation, especially for
plasmids with a
low copy number, so eventually descendants of the host cell contain only
one or the
other plasmid.
Virulence Plasmids
Virulence plasmids contain genes that make their bacterial hosts more
virulent to their
hosts. A familiar example involves the bacterium E. coli, which inhabits
the human
large intestine. Certain strains of E. coli contain plasmids whose genes
make the
E. coli synthesize toxins that cause diarrhea. These "enterotoxigenic
strains" of
E. coli are probably an important cause of diarrhea among travellers.
More seriously,
in developing countries, diarrhea is one of the principal causes of death
among those
under five.
"Vibrio cholerae", the cause of cholera, is a bacterium whose
genes code for a
diarrhea-causing toxin. The DNA of these genes is closely related to the
DNA of
certain virulence plasmids infecting E. coli - so closely that there is
almost
certainly a common ancestor. For example, Vibrio cholerae could have evolved
from an earlier bacterium by permanently integrating the DNA from a virulence
plasmid into its genome.
Strains of bacteria and viruses often become less virulent as they coevolve
with their
hosts. Thus one may wonder what evolutionary advantage a virulence plasmid
could
confer to the bacteria containing it. In the case of bacteria causing
diarrhea, there
is an obvious possibility: diarrhea can serve as a mechanism for spreading
the
bacteria - and their plasmids - that cause it!
Metabolic Plasmids
Metabolic plasmids contain genes that let their bacterial hosts metabolize
or
degrade otherwise indigestible or toxic chemicals. For example, the bacterium
Pseudomonas putida is able to grow on a wide range of organic compounds
that
are toxic to most bacteria, including toluene, octane, camphor, napthalene
and
nicotinic acid! It does this with the help of genes contained by metabolic
plasmids
called TOL, OCT, CAM, NAH and NIC plasmids.
It's worth noting that some of these chemicals are secreted by plants
as part of a
defense against bacteria. Thus we probably have a kind of natural chemical
arms
race going on here. Other metabolic plasmids allow bacteria to degrade
herbicides
like 2,4-D, as well as certain detergents! People are investigating the
use of such
plasmids to help biodegrade pollution.
Tumor-Causing Plasmids
"Crown gall" is a cancer of plants caused by a bacterium known
as Agrobacterium
tumefaciens. But actually, the disease is caused by a plasmid having this
bacterium
as its host! When the plasmid passes from the bacterium to the cells of
infected fruit
trees, some of the genes contained in the plasmid cause tumors. Do these
tumors
help spread the bacteria to other trees?
Cryptic Plasmids
Cryptic plasmids are plasmids that have no known effect on their hosts.
How much
of this is our ignorance, and to what extent is being truly "cryptic"
a successful strategy?
Cosmids
Cosmids are man-made circular loops of DNA containing plasmid DNA together
with an arbitrary sequence of up to 45,000 base pairs of DNA. They are
constructed
by recombinant DNA techniques and then packaged in lambda phage protein
coats.
They are used to transfer genes to bacteria.
The lambda phage is a virus that specializes in invading bacteria such
as E. coli.
In nature, its protein coat latches onto the bacterial cell membrane and
injects the
phage DNA into the bacterium. Biotechnologists have taken advantage of
this
by using the lambda phage protein coat to inject a cosmid into the bacterium!
Once inside, the cosmid replicates like a plasmid and, like a plasmid,
integrates
its DNA into the genome of the bacterium.
Phasmids
Phasmids are man-made linear DNA molecules whose ends are sequences taken
from the lambda phage, while the middle is a sequence taken from a plasmid,
together with a sequence of whatever DNA one wants. Like cosmids, they
are
constructed by recombinant DNA techniques and packaged in lambda phage
protein coats, and used to transfer genes to bacteria. However, both the
lambda
phage and plasmid replication functions are intact. In particular, they
contain the
lambda phage genes for "lysis", the process whereby a virus
dissolves the cell
membrane of its host. Depending on the conditions, the phasmid can act
either
like a phage or a plasmid - hence its name.
--------------------------------------------------------------------------------
Transposons
Transposons, or "transposable elements", are sequences of DNA
that move within
their host's genome from one position to another. They were first discovered
in the
1940s by Barbara McClintock, who later won the Nobel prize for this work.
They
exist in all known organisms, often in large quantities. Their main "function"
appears
to be simply their own self-replication, rather than any benefit to the
host, or even
any direct effect whatsoever on the host phenotype. For this reason, people
sometimes
refer to transposons as "selfish DNA".
In addition to transposons, there is plenty of other DNA in our chromosomes
that
doesn't seem to code for proteins. This is sometimes called "junk
DNA". It comes
in various distinct forms, such as "introns", "satellite
DNA", and "pseudogenes".
In fact, junk DNA makes up about 97% of the human genome! Clearly despite
its
derogatory name, it's worth understanding and potentially very important.
However,
since transposons are the most "organism-like" of junk DNA,
I will only talk about
them here.
There is a fair amount of genetic evidence that transposons spread "horizontally"
between sexually isolated species in addition to being "vertically"
passed down
the evolutionary tree. However, the mechanisms of this horizontal transmission
are poorly understood. One interesting fact is that certain viruses, the
baculoviruses, can pick up and accomodate transposons from their hosts.
They have been proposed as a possible mechanism for horizontal transmission
of transposons.
The two main classes of transposons are:
Retrotransposons
and
DNA Transposons
The best book on transposons seems to be "Dynamics and Evolution
of
Transposable Elements", by Pierre Capy, Claude Bazin, Dominique Higuet,
and Thierry Langin [CBHL]. In this book, retrotransposons are called
"Class I elements",
while DNA transposons are called "Class II elements". They also
discuss
"Class III elements". This seems to be a grab-bag consisting
of transposons
that don't clearly fit into the other two categories. Examples include
the "Foldback"
elements in fruit flies, the "Tu" elements in sea urchins, and
"MITEs",
or "miniature inverted repeat transposable elements", which
are found
mainly in plants and fungi.
Retrotransposons
Retrotransposons copy themselves from one location in the host genome
to
another using an RNA intermediate, with the help of reverse transcription
from RNA to DNA.
A rough classification of retrotransposons divides them as follows:
LTR (long terminal repeat) retrotransposons
non-LTR retrotransposons
LINEs (long interspersed nuclear elements)
SINEs (short interspersed nuclear elements)
LTR retrotransposons are are 5000-9000 base pairs long and have
" long terminal direct repeats" - repeating sequences of base
pairs
at both ends. Between these are the genes needed for transposition,
which code for enzymes like reverse transcriptase
(which copies RNA into DNA), integrase (which integrates the DNA
into the host chromosome), and so on. In all these respects,
LTR retrotransposons are very similar to retroviruses.
The most important difference is that retrotransposons do not code
for the proteins forming the viral protein coat. There seems to be
some debate as to whether retrotransposons are retroviruses that
have somehow lost their ability to code for a protein coat, or whether
retroviruses are retrotransposons that have somehow gained this ability.
Of course, the two possibilities aren't mutually exclusive!
As the name suggests, non-LTR retrotransposons lack terminal repeats.
They have been divided into LINEs and SINEs. LINEs have a characteristic
adenosine-rich sequence at one end, and are generally 5000-8000 base pairs
long, though truncated versions are common. They code for various enzymes
such as reverse transcriptase and RNase. The genomes of higher animals
and plants may have over 10,000 copies of LINEs. In fact, at least 15
percent
of the human genome consists of LINEs!
SINEs are usually shorter than 500 base pairs. The source of the enzymes
needed for the mobility of SINEs is not yet known - but perhaps it is
LINEs!
Higher animals and plants may have over 100,000 copies of SINEs.
DNA transposons
DNA transposons mainly move using a cut-and-paste mechanism: they code
for an enzyme called a "transposase" that catalyzes a process
in which the
transposon DNA is excised and reinserted elsewhere in the host genome.
Thus RNA and reverse transcriptase plays no role in their life cycle.
--------------------------------------------------------------------------------
Prions
Prions are small, proteinaceous infectious particles that contain no detectable
nucleic acid of any form, but are transmissible among certain animals,
where they cause fatal brain diseases. These particles are rod-shaped,
about 165 nanometers long and about 11 nanometers in diameter, and
they consist largely of a protein called PrPSc, having molecular
weight 33,000-35,000. They are able to resist inactivation by boiling,
acid (pH 3-7), ultraviolet radiation (254 nm), formaldehyde, and
nucleases! They can be inactivated by boiling in detergents, alkali (pH
> 10),
autoclaving at 132 degrees centigrade for over 2 hours, and denaturing
organic solvents such as phenol.
Stanley Prusiner won the Nobel prize for medicine in 1997 for his work
on prions.
His theory is that prions are a modified form of a protein naturally occuring
in
the brain (PrP), and that this modified form can arise from a cell mutation,
but
then spread by means of a kind of autocatalyzed chain reaction. This theory
was initially very controversial, because all other self-reproducing biological
entities appear to contain RNA or DNA. There are still many doubters.
In the earlier literature prions are sometimes called "slow viruses",
because of their slow effect. However, no virus has ever been associated
with prion diseases.
Prions have recently received a lot of publicity as the cause of
"mad cow disease", technically known as bovine spongiform encephalopathy.
Starting in the mid-1980s, this disease infected thousands of cattle in
England,
in part because they were being fed offal containing nerve tissue from
sheep
infected with a prion-caused disease called "scrapie". People
got worried that
eating meat from cows with bovine spongiform encephalopathy could cause
a prion-induced brain disease in people. This caused an enormous uproar.
There are already a number of prion-induced brain diseases in people,
such
as Creutzfeldt-Jakob disease (which occurs spontaneously in about one
in a
million people) and kuru (transmitted by means of cannibalism among the
Fore tribe in New Guinea). There are also prion-induced brain diseases
in mink,
cats, deer and moose.
--------------------------------------------------------------------------------
References
Here are some good books to read about this stuff:
[B] Plasmids, by Paul Broda, W. H. Freeman, San Francisco, 1979.
[CBHL] Dynamics and Evolution of Transposable Elements, by Pierre Capy,
Claude Bazin, Dominique Higuet, and Thierry Langin, Landes Bioscience,
1998.
[CT] Principles of Bacteriology, Virology and Immunity, vol. 4: Virology,
edited by L. H. Collier and M. C. Timbury, 8th edition, Decker, 1990.
[D] The Viroids, edited by Theodore Otto Diener, Academic Press, 1985.
[E] Plasmids of Eukaryotes: Fundamental and Applications, K. Esser et
al,
Springer-Verlag, New York, 1986.
[F] Virology, 2 volumes, edited by Bernard N. Fields, David M. Knipe
and
Peter M. Howley, Lippincott-Raven Publishers, 3rd edition, 1996.
[H] Bacterial Plasmids, Kimber Hardy, American Society for Microbiology,
Washington D.C., 1986.
[MM] Subviral Pathogens of Plants and Animals: Viroids and Prions,
edited by K. Maramorosch and J. J. McKelvey, Jr.., Plenum Press, 1987.
[Ma1] Viroids and Satellites: Molecular Parasites at the Frontier of
Life,
edited by Karl Maramarosch, CRC Press, 1991.
[Ma2] The Atlas of Insect and Plant Viruses: Including Mycoplasmaviruses
and Viroids, edited by Karl Maramorosch, Academic Press, 1977.
[Mo] The Evolutionary Biology of Viruses, edited by Stephen S. Morse,
Raven Press, 1994.
[R] Plant Infectious Agents: Viruses, Viroids, Virusoids, and Satellites,
edited by Hugh D. Robertson et al., Cold Spring Harbor Laboratory, 1983.
THE ROBERT CATHEY RESEARCH SOURCE
http://www.navi.net/~rsc
--------------------------------------------------------------------------------
[Originally Published in: U.S. Psychotronics Proceedings, 1993; ]
Royal Rife Revisited:
Reconstruction of the Original Rife Ray Tube
by
David M. Tumey and William H. Sheline
Abstract -- In this paper the authors relate information obtained from
over
three years of work spent researching and reconstructing a working
replica of Royal Rife's original Ray Tube apparatus. A description of
Rife's
discovery of the pleomorphic nature of microorganisms is given along with
details of how this led him to invent a revolutionary non-invasive pathogen
devitilization technique. Also discussed with limited detail was Rife's
development of his extraordinary microscopes. The authors attempt to give
the reader new insights into this exciting, readily available technology.
Finally, the authors describe the design and fabrication of a complete
and working beam tube system, constructed with antique and surplus
electronic components. This paper attempts to provide enough information
so that all can understand what it was Rife was attempting to accomplish,
how Rife's machines worked, and how similar machines might be
manufactured today. Also, a list of the original known Mortal Oscillatory
Rates (MORs) is provided. No claims for the use of this device in healing
human subjects are made.
Introduction -- Royal Raymond Rife (1888 - 1971) was an accomplished
scientist and microbiologist who developed an optical microscope that
could provide magnifications and resolutions heretofore unheard of.
He was able, through special quartz optics and a creative optical
heterodyning technique, to obtain these resolutions even though they
surpassed the theoretical limits of ordinary visible light microscopy.
Theoretically, the wavelength of the source illumination is the limiting
factor in achievable resolution. It is not possible to image something
smaller than the wavelength of the microscope's light source. That is
why electron microscopes (with far shorter wavelength) can be used
today to image extremely small objects. The major difference between
visible light and electron microscopy is that, by its nature, electron
microscopes destroy the microorganisms while viewing them. Rife's
major advantage was that he could observe them in their natural state.
His most powerful instrument is said to be the Universal Microscope
which had a magnification of 61,000X and a resolution of 30,000 diameters.
Compare this with today's state-of-the-art light source microscopes
which are limited to approximately 5,000 diameters.
Rife began his work with the microscopes in the early 1920's and it was
from these original developments that he would make many of his
revolutionary discoveries. It is argued that Rife was the first person
to empirically prove that virus and bacteria are pleomorphic forms.
Pleomorphism is the phenomenon by which one distinct life form
mutates into another. Rife basically classified pathogenic bacteria
into 10 individual groups. Rife demonstrated that any organism
within its group could be transformed morphologically into any
other organism within the 10 groups by carefully altering the media
in which it was cultured. Of course this discovery contradicts modern
microbiology which teaches that a bacteria's morphology is fixed and
unchangeable.
Rife also discovered techniques for successfully culturing cancer virus. This virus he identified as BX and it was noted that the viruses refracted a purplish red color with a monochromatic beam under his microscope. In fact, Rife discovered that each organism depending on its state would refract unique spectra and have distinct coloration. By the late 1920's and early 30's, Rife had discovered that by irradiating these pathogenic microorganisms with specific frequencies known as MORs for Mortal Oscillatory Rates, he could cause them to devitalize either by interrupting normal cytologic function or by inducing them to mutate into a non-pathogenic form.
The instrumentation involved in this irradiation process has been the subject of a great deal of controversy over the past 50 years. After researching books, films, articles and notes, the authors have concluded that Rife irradiated his pathogenic entities with a modulated radio frequency produced by a sophisticated RF plasma discharge. Rife utilized a radio frequency generator that produced between 100 and 150 Watts of power with a carrier frequency between 3.1 Mhz and 35 Mhz. The output from the generator was connected through suitable impedance matching circuitry to a plasma discharge tube with one or more noble gases. It is believed that Helium was the primary gas used although many researchers cite Argon or an Argon mix as the choice ingredient. Further, Rife utilized a standard dial-type vacuum tube audio frequency generator as the modulation source for his radio frequency transmitter. The modulation signal was a square wave and it is assumed he chose this waveform because of its high harmonic content and broad spectral contributions.
Rife obtained the original MORs through a painstaking method of tuning the dial of the audio frequency generator while observing the sample pathogen under his microscope. When a frequency was discovered that demonstrated the ability to devitilize a particular microorganism, its dial position was duly noted and marked. The actual frequencies were determined later after the experimental trials. By the mid 1950s the verified original MOR frequency list included 15 different bacteria and viruses. Regardless of what other researchers have said, the authors believe that these 15 frequencies represented the complete list. The following is a listing of these known MORs as compiled by Dr. Robert P. Stafford, M.D. a physician who worked with an original Rife machine from 1957 to 1963:
Microorganism Frequency in Hertz
------------- ------------------
Tetanus 120
Treponema 660
Gonorrhea 712
Staphlococci 728
Pneumococci 776
Streptothrix (fungus) 784
Streptococci 880
Typhoid Bacteria 712
Typhoid Virus 1862
Bacillus Coli Rod Form 800
Bacillus Coli Virus 1552
Tuberculosis Rod Form 803
Tuberculosis Virus 1552 (same as B-Coli)
Sarcoma (all forms) 2008
Carcinoma (all forms) 2128
Dr. Stafford who is still living in Dayton, Ohio, independently verified
some
of Rife's work. Dr. Stafford conducted a rat study with the assistance
of
Dr. Robert Zipf, M.D., who at the time was the Director of Medical
Research at Miami Valley Hospital and in addition, the Montgomery
County Coroner. Chloroleukemic Sprague-Dawley rats were utilized in
the experiment. Although it is beyond the scope of this paper to discuss
the study results in detail, the encapsulated summary is as follows:
Ten suckling rats were injected with standard doses of rat leukemic
whole blood. Of the seven rats which were inoculated and treated with
the Rife equipment, three survived without symptom. Four of these rats
died. However, the average time to death was 50.5 days as compared to
the group of three non-treated rats which had an average death time of
43.6 days. In addition, all the non-treated rats died. Clearly, even with
the four 'failed-cures', in the group of treated rats the Chloroma was
favorably impressed.
Details of the Original Tube -- In the "Cancer Cure that Worked",
Barry Lynes includes a quote penned by Rife himself, which describes
the principles of the original Ray Tubes. Also, photographs of original
tubes can be found in "The Rife Way III" by Mark Simpson. Figure
1
details the component arrangement of the early units (prior to the formation
of Beam Ray Corp.). As described in the introduction, a standard frequency
generator (a) was utilized as the square wave modulation source.
The individual MORs were selected for broadcast via this instrument.
The function generator was connected to the radio frequency generator
(b). The square wave signal from the frequency generator was employed
in a screen-grid modulation arrangement with the final amplifier of the
radio frequency generator. The RF generator incorporated circuitry bearing
resemblance to a standard radio transmitter and had a single stage crystal
controlled oscillator connected to a class-C amplifier stage. Most likely,
the oscillator used either a 6AG7 or 6V6 vacuum tube, while the output
was obtained from a pair of standard RF transmitter tubes. RCA designed
807's were introduced in the later models (1940's).
The output from the amplifier was connected to an impedance matching
network (c) designed to maximize the transfer of RF power from the
generator to the gas plasma. As with any transmitter, maximum
power transfer occurs when the impedance of the generator is the
same as the impedance of the load. The major problem with matching
to a plasma is that the plasma's impedance is dynamic and highly
non-linear. Rife must have been faced with a formidable challenge
having to address the many variables involved in controlling an RF
plasma. The original plasma tubes were modified X-ray tubes (d).
The tube elements were left intact, however, he tube was refilled
with one or more noble gases. When the plasma discharge occurred,
the tube glowed purplish blue as attested to by Dr. Stafford,
John Crane (Rife's research assistant from 1950 - 1971) and others.
Later, Rife had several tubes fabricated from scratch. It is not known
precisely how many of this genus of tube were manufactured.
Although records are difficult to accurately verify, it is generally
thought
that patients were treated every third day by exposing them to each frequency
on the list for three to four minutes. Normally, only a subset of frequencies
were used, a popular list being 728, 784, 880, 2008, and 2128 Hertz.
While the modulation frequencies were being broadcast to the patient,
the discharge tube was moved over the body of the subject.
It was determined that this kind of motion greatly enhanced the
experimental effects. Also, an additional improvement was obtained
when the MOR frequencies were dithered +/- 10 Hertz. Some believe
that in combination with the MOR rates, the RF generator was gated
on and off at a 4 hertz rate. Early film footage of the Rife tube in operation
does tend to support this conclusion as the tube is seen to flash repetitively.

Building a working Beam Tube -- To date, the authors have constructed
a total
of four Rife units. Two units were fabricated around modified Heathkit
transmitters and delivered 100 and 250 Watts respectively. Two additional
units were designed from scratch, one utilizing a single 6DQ6 tube as
both
an oscillator and amplifier. This unit produces approximately 50 Watts
of
input power. The second and favorite system utilizes a 6AG7 crystal controlled
oscillator stage driving a single 807 amplifier. This unit produces about
75
Watts and is ideal for work with RF plasmas. The 807 is an excellent RF
amplifier tube in that it is extremely forgiving and will deliver the
full rated
75 Watts output with only 220 milliwatts of grid drive. Each of the units
employs a solid state modulator in the cathode circuit of the amplifier
stage. This cathode arrangement acheives 100% modulation more
readily than the screen-grid type originally employed by Rife.
The schematic for this unit can be seen in Figure 2.
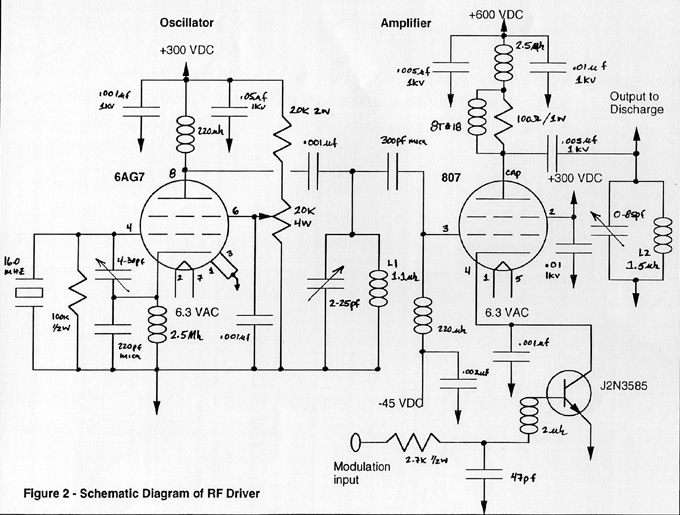
RF Driver, click here to view larger format (121K)
The most difficult part of the reconstruction effort was obtaining the glasswork for the plasma discharge tube. Several models were constructed using estimates obtained from descriptions and photographs of original tubes. In addition, physical dimensions were taken from several X-ray tubes designed in the 1940's. Tube fabrication was fabulously expensive and required many tedious hours spent measuring the effective impedance of various gases under varying pressures and power levels. A tremendous amount of time was spent researching plasma chemistry in an attempt to understand the physics behind radio frequency plasma formation and control.
If fabricating a replica tube sounds cost prohibitive, please note that a suitable discharge tube can be obtained from a neon sign manufacturer. This less expensive option is highly recommended for a novice experimenter. Have a local glass shop make a 16-19" straight tube and ask them to fill it with argon or helium. This type or discharge tube will operate quite reliably and require no periodic maintenance. Detailed discussions on RF plasmas are beyond the scope of this paper, so it shall be left to the reader to pursue further information on the subject. Most technical libraries will have several informative selections pertaining to RF plasma chemistry.
Referring to Figure 2, starting with the oscillator stage, a 6AG7 tube was chosen because its plate load resistance is close to the input impedance of the grid of the 807. This serves to maximize power transfer from the oscillator to the amplifier stage. These impedances can be calculated by using equations and tube data found in older Amateur Radio Handbooks. The 807 requires -45 volts on the grid along with 3.5 milliamps of grid drive for proper excitation. The screen voltage nominally is 300 and the plate can be run at 600 d.c. volts. The driving characteristics of the 807 present an input impedance of approximately 11,000 ohms. A standard tuned-plate tuned-grid crystal oscillator circuit was taken from the handbook along with the recommended component values. The crystal used was cut for 16.0 Mhz. The screen grid of the 6AG7 was fed through a variable 20,000 ohm potentiometer so that the necessary 3.5 milliamps of the grid drive could be carefully adjusted. This control prevents the oscillator from overdriving the amplifier tube. The output from this stage was coupled to the 807 with a simple series capacitor. A fixed regulated supply was chosen over a grid-leak configuration for the grid bias circuit because if the oscillator were to fail, the 807 could be severely damaged. The fixed supply serves as a protective bias for the amplifier.
The plasma discharge tube presents an approximate impedance of 2000 ohms at 16.0 Mhz. With the 807 operating with 600 volts on the plate and 150 miliamps of plate current, the output impedance is 4000 ohms, therefore, the discharge tube is tapped down on the output tank coil enabling it to present a higher impedance to the amplifier tube via the transformer action of the circuit. Since the load impedance is matched at 4000 ohms, and the desired loaded Q of the circuit is approximately 15, the tank inductor should exhibit a reactance of 266 ohms at the carrier frequency. This equates to an inductance of 2.65 microhenrys at the 16.0 Mhz operating frequency. A variable capacitor with a range of 5 to 50 picofarads was chosen to resonate with this coil.
Since the discharge tube is highly non-linear and has reactive components contributing to its overall impedance, proper adjustment of the variable capacitor allows almost complete cancellation of these reactive components. When the tank circuit is tuned to resonance, the load impedance appears to be purely resistive. This enables the power to be completely transferred from the amplifier to the discharge and be dissipated as radiant energy.
Operation and adjustment of the RF driver is fairly simple. Power is first applied to the filaments of the tubes and the tubes are allowed to warm up for several minutes. After the tubes have reached nominal operating temperature, with the amplifier cathode circuit open, high voltage is applied. The drive tuning and drive level of the oscillator stage are adjusted to produce 3.5 milliamps of grid drive for the 807. Next, the cathode circuit is closed and the plate tank capacitor is adjusted to obtain the brightest discharge, which should also require the least amount of plate current, since the circuit will be at resonance with all reactances cancelled. The tap for the discharge tube on the tank coil is adjusted to produce the desired 150 milliamps of d.c. plate current. Modulation can now be accomplished by connecting a suitable square wave function generator to the modulation input. The square waves should have a peak to peak level between 10 and 15 volts. A model B&K-3011 function generator will serve nicely in this capacity.
Conclusion -- Now that the Rife Beam Tube technology is fairly well understood and can be duplicated with a high degree of confidence, the next logical step might be to repeat the laboratory studies originally performed by Dr. Stafford et al. Also, a concerted effort should be undertaken towards recreating a working Rife type microscope. Much has been written recently about dark-field microscopy and advances made in Canada by Gaston Naessens and others. Their efforts need to be supported if this technology is ever to be fully realized.
This paper did not discuss the "Rife Machines" recently popularized by a number of commercial equipment manufactures. This type of unit normally incorporates either hand-held electrodes or foot plates. It is believed that these devices emerged in the 1950s and are more a product of John Crane than Royal Rife. Rife's involvement in the development of this genre of machine is at best unclear. Although the authors have heard of positive experimental results obtained with the electrode units, they feel that the original Beam Tube system invented by Rife himself offers the best hope as a viable treatment modality. The authors would raise a caution to any individual utilizing any device like this as part of a therapeutic regimen.
For additional information on reconstructing a working Rife Beam Tube, contact the authors through the Phoenix chapter of the USPA.
[Update: The authors are preparing a second paper, and may be contacted at]:
Open Sesame
601 W. Leffels Lane, Suite J
Springfield, Ohio 45506
937-290-6758 voice mail
References:
"The Cancer Cure that Worked", Lynes, Barry, (1987), pp. 72-73.
"The Rife Way III", Simpson, Mark A., (1991), pp. 2-13.
"The New Microscopes", Seidel, R.E. and Winter, M. Elizabeth, Journal of the Franklin Institute, February, 1944.
"Observations on Bacillus and Typhosus in its Filtrable State", Kendall, Arthur and Rife, Royal, California and Western Medicine, December 1931.
"Observations with the Rife Microscope of Filter-Passing Forms of Microorganisms", Science, August 26, 1932.
"Techniques and Applications of Plasma Chemistry", Hollahan and Bell, (1974), pp. 393-399.
"The Radio Amateur's Handbook", American Radio Relay League, Newington, Conn.
"RCA Power Circuits -- D.C. to Microwave", Radio Corporation of America, Harrison, N.J.
"Care and Feeding of Power Grid Tubes", Eimac Division of Varian, San Carlos, Ca., (1967).
"RCA Transmitting Tubes", Radio Corporation of America, Harrison, N.J.
"The Semiconductor Data Library", Motorola Semiconductor Products, Inc., Pheonix, Ariz.
--------------------------------------------------------------------------------
(Current Document Location: http://www.navi.net/~rsc/riferev.htm)
RETURN TO INDEX
The following is a listing of known MORs (mortal oscillatory rates) as compiled by Dr. Robert P. Stafford, M.D. a physician who worked with an original Rife machine from 1957 to 1963. They are believed to be the killing frequencies Royal Rife himself discovered and verified with his microscope. (from http://www.alteredspace.com/~rsc/riferev.htm)
Original Rife frequencies
Tetanus 120 (An acute, often fatal infectious disease caused by the anaerobic, spore forming bacillus Clostridium tetani.)
Syphilis 660 (Caused by Treponema pallidum, a helical, tightly coiled, motile spirochete, a helical to sinusoidal bacterium. Mechanisms of T. pallidum pathogenesis are poorly understood. Existing diagnostic tests for syphilus are sub-optimal, and no vaccine against T. pallidum is available. The subspecies of T. pallidum cause syphilis, yaws, nonvenereal endemic syphilis or pinta.)
Gonorrhoea 712 (A gram-negative bacteria, Neisseria gonorrhoea, causes this sexual disease and primarily affects columnar epithelium in genital mucosal surfaces of the urethra, accessory ducts and gland, as well as endocervix. If contaminated fingers rub the eye then conjunctivitis can result.)
Staphlococcus 728 (Genus of nonmotile gram-positive bacteria that are found in clusters and that produce important exotoxins. Staphylococcus aureus (Staphylococcus pyogenes) is pyogenic, an opportunistic pathogen and responsible for a range of infections including severe sepsis, pneumonia, endocarditis and soft tissue infections.)
Pneumococcus 776 (Gram-positive pyogenic organisms about 1m diameter, usually encapsulated, closely related to streptococcus, associated with diseases of the lung. Pneumococcus is an important cause of serious infections in the first three months of life. These infections are unlikely to be prevented by the currently available infant immunization strategies. One potential approach to prevention of pneumococcal disease in early infancy is immunization of pregnant women.)
Streptococcus 880 (A genus of bacteria that are gram-positive cocci, often occurring in chains of varying length. Some pathogenic species produce exotoxins. In man, streptococcal species are responsible for numerous infections such as scarlet fever, tonsillitis, erysipelas (skin infection), endocarditis, rheumatic fever, glomerulonephritis, impetigo, pneumonia, meningitis, pharyngitis, lymphadenitis and wound infections. Streptococcus pneumoniae is the main culprit in lobar-pneumonia and broncho-pneumonia. Streptococcus pneumoniae has been known for more than 100 years as the most important bacterial pathogen of the respiratory tract in adults and children. In recent years, the pneumococcus has begun to exhibit increasing resistance to antimicrobial agents.)
Typhoid Bacteria 712 (Typhoid is an infectious febrile illness usually spread by contamination of food, milk or water supplies with bacteria Salmonella typhi. This is not to be confused with Salmonella typhimurium which is the cause of salmonella food poisoning.)
Typhoid Virus 1862
Bacillus Coli Rod Form 800 (Most probably Escherichia coli, the archetypal bacterium for biochemists, used very extensively in experimental work. A rod shaped gram-negative bacillus (0.5 x 3-5 m) abundant in the large intestine (colon) of mammals at about .1% of the total. E. coli, along with other species of bacteria, provide us with Vitamin K and B-complex vitamins. But a rare strain of this bacteria, E. coli O157:H7, is responsible for food poisoning which causes bleeding of the intestines which can be fatal.)
Bacillus Coli Virus 1552
Tuberculosis Rod Form 803 (Tuberculosis, an infectious bacterial disease caused by Mycobacterium tuberculosis, is characterized by inflammatory infiltrations, formation of tubercles (solid elevations of skin or mucous membranes), tissue death, abscesses, formation of fibrous tissue, and calcification of tissue. Infection is transmitted from infected people, cows, or contaminated milk. Presently the worlds leading killer. It usually occurs as pneumonia, but TB can also occur in the brain, back, knee, lymph nodes, or other organs and bones.)
Tuberculosis Virus 1552
Sarcoma cancer (all forms) 2008 (A form of cancer that arises in the supportive tissues such as bone, cartilage, fat or muscle.)
Carcinoma cancer (all forms) 2128 (A malignant new growth that arises from epithelium, found in skin or, more commonly, the lining of body organs, for example: breast, prostate, lung, stomach or bowel. Carcinomas tend to infiltrate into adjacent tissue and spread (metastasize) to distant organs, for example: to bone, liver, lung or the brain.)
Streptothrix 784 (A genus of bacilli occurring of the form of long, smooth and apparently branched threads, either straight or twisted. Streptothrix is a synonym for Actinomyces israelii. This species is a gram-positive, cast-forming, non–acid-fast, non–spore-forming anaerobic bacillus that is difficult to isolate and identify. Its filamentous growth and mycelialike colonies have a striking resemblance to fungi. They are soil organisms. It can cause the eye diseases Canaliculitis and Keratitis.)
Also, frequencies for leprosy, polio, cholera, actinomycosis, glanders, bubonic plague, anthrax, influenza, herpes, cataracts, glaucoma, colitis, sinus, ulcers were discovered by Rife but we don't have any direct records from him on these frequencies.
--------------------------------------------------------------------------------
(Definitions in parentheses are mostly from the on-line medical dictionary at http://cancerweb.ncl.ac.uk/omd/index.html and other medical web sites.)
Rife stated they had narrowed the actual distinct number of groups of
pathogenic bacteria to 10. In his 1953 book, Rife commented on this:
"We have classified the entire category of pathogenic bacteria into
10 individual groups. Any organism within its group can be readily changed
to any other organism within the ten groups depending upon the media with
which it is fed and grown. For example, with a pure culture of bacillus
coli, by altering the media as little as two parts per million by volume,
we can change that micro-organism in 36 hours to a bacillus typhosis showing
every known laboratory test even to the Widal reaction. Further controlled
alterations of the media will end up with the virus of poliomyelitis or
tuberculosis or cancer as desired, and then, if you please, alter the
media again and change the micro-organism back to bacillus coli."
Rife contended certain conclusions escaped earlier researchers simply because they lacked the evidence of their eyes in seeing these forms develop from a single entity: pleomorphism. They require a power of magnification and resolution beyond the typical 2,000 power instrument.
Rife's work suggested that the wide array of disease bacterium were merely differentiation phases in a life-cyle of an as of yet undetermined entity. Researcher Gaston Naessens has verified many of Rife's findings, and has delineated 16 phases of change of what Rife called the premodal identity, which Naessens calls "somatids".
The Rife frequency instrument kills the "normal" carcinoma cancer cell by rupturing the thousands of BX cancer viruses they contain and thereby dumping the BX cancer virus contents into the cancer cell cytoplasm. This BX cancer virus as Rife named it in 1931 is not a virus by the normal standard usage of the term today. Rife based his definition on the fact that the BX cancer virus could pass through the finest Berkefeld porcelain filter of the time (000 filter). The BX cancer virus is ovoid in shape, .066 microns along the major axis and .05 microns along the minor axis. It is motile, driven by a proton transport flagella the same as its bacterial parent, the E-coli bacteria. When the BX cancer virus is ruptured it spills out its genome, ribosomes, RNA, enzymes, and various proteins. When thousands of these ruptures occur all at once in a carcinoma cancer cell the results are fatal to the cancer cell. A similar situation occurs in the sarcoma cancer cell when the BY cancer viruses are all disintegrated at once. The BY cancer virus is another form of the BX cancer virus which Rife found caused sarcoma cancer after it had been exposed to prolonged ultraviolet light exposure.
Royal Rife's Laboratory Research on Bacillus "X" cancer virus
The BX was isolated from ten different cases of breast carcinoma by Dr.
Royal Raymond Rife at the Rife Research Laboratory in SanDiego (Point
Loma), California. It was carried through forty-four transplants on "K"
media in all ten instances.
The technique used in the isolation of this organism is in brief as follows;
blocks of tissue, taken under the most sterile
conditions, were transferred into "K" media (previously examined
for sterility). These were then placed under the direct
influence of an argon filled gas tube working under five thousand volts
for twenty four hours, then were placed in water baths
with two inches of vacuum, and incubated at 37.5°C. At this time the
delicate shine of growth is noticeable. From this point on
as many as desired transplants can be made without repeating the foregoing
operations.
The BX is a filterable virus, which filters through the W Berkfeld filter.
It is a small ovoid granule, highly plastic, and visible only
with monochromatic light. The angle of refraction is 12 3/10°, and
the color by chemical refraction is purple red. The length of
the organism is 1/15 u, and its breadth 1/20 u. It carries an attraction
to the cathode pole. Its death rate in milliamperes is 175
DC. The X-ray and Infrared have no influence on the organism, but the
Ultraviolet ray slows up its motility. The thermal death
point is 42°C for 24 hours, the filament voltage is 10, and the filament
amperage is 86. The plate voltage is 928, and its
unmodulated electronic oscillatory rate is 11,780,000 cycles per second.
The wavelength of super regeneration of Audion tube is 17 6/10 meters.
An inoculating serum was prepared by combining in a mixture, the transplants from the ten original growths with a (?) to 1 dilution of normal saline solution.
(The symbol (?) indicates that the text was cropped out in the photocopy.)
On Aug. 3, 1933 1/10 cc of the above serum was inoculated into the breasts
of two sets of white rats; one set consisting of
two pregnant females with one control, and the other of two young females
and one control. The animals had been kept in
quarantine for a period of ninety days and were normal in every respect
at time of inoculation. Seven days later the inoculated
rats developed lesions (superficial) in the thyroid region and on the
shoulders. These lesions varied in size and severity on
succeeding days. The controls remained normal. On Aug. 21, the control
of the pregnant female set gave birth to two young;
one died. The delivery of the inoculated pregnant females was still delayed,
and the temperatures of all the inoculated animals
rose from 1 to 1 1/2° F. The lesions increased in area and density,
and one in particular was decidedly elevated. On Aug. 22,
one of the infected rats presented 5 young, and the other until this day
has remained barren, the swelling of the abdomen which
evidently was occupied by the young having gone down and returned to normal.
In the offspring of the infected mother, two
developed the identical type of lesions on the surface of the thyroid
region. One of these grew otherwise normally and in the
other the growth was stunted. The latter developed a severe growth on
the upper portion of the right side of the jaw, which
consumed most of the normal tissue. The teeth were badly malformed, and
grew very long, curving down and deep into the
throat. These were shortened by surgical operation. During this entire
period the controls remained normal.
On Aug. 28, a set of male rats consisting of the same number was inoculated
as in the females. The same type of epidermal foci
developed, the control remaining normal. On Sept. 5, one of the males
was posted and revealed no pathology. A lesion was
excised from the shoulder of the other inoculated male. (*) On Sept. 14,
the Bacillus X was recovered and identified in the
media. The lesions on all the inoculated rats vary in size and density
from day to day and in some cases clear up and break out
in other portions of the epidermis.
(*) Tissue placed in "K" media and run through the original method of technique.
It has been demonstrated by experiment that the BX exists in two cycles,
which may be classified as forms A and B. Form A
applies to BX in its ultra filterable cycle. In this stage the organisms
theoretically exist in malignant tissue. Examination of the
fresh filtrate preparation of malignant tissue under 20,000X magnification,
using any known system of illumination, fails to reveal
the presence of living bodies. However, after a special method of cultivation,
involving the use of the argon ray and vacuum
conditions, the aforementioned filtrate in "K" medium contains
a swarming myriad of the visible cycle, form B. The BX in this
form may be seen under 8-11,000X magnification, (using monochromatic illumination)
as a highly plastic ovoid granule, purple
red in color.
Since experiments show that the Bacillus X in form A exists in malignant
tissue, it is theoretically possible to change its cycle to
form B by application of the argon ray and vacuum conditions. After the
cycle change has been accomplished (in theory), the
application of the oscillative ray at a cycles per second vibration of
11,780,000 should completely destroy the BX in the
malignant tissue.
(note: the frequency listed here is the base frequency that undergoes modification by the audio frequency that isn't listed here which is believed to be 2127 or 2008.)
The Forty Year Legacy of Tainted Polio Vaccine
by
Harold Stearley
In the late 1940's and early 1950's the polio virus was taking a savage toll on the American public. Thousands of children and adults were crippled or killed. In 1955, Jonas Salk performed a medical miracle when he discovered how to mass produce polio vaccine by growing it on the kidneys of rhesus monkeys. While there is no question that thousands were saved from the ravages of polio by the Salk vaccine, by 1960 a problem had surfaced -- a problem which would come back to haunt the nation some forty years later.
The complication researchers had isolated in 1960 was a viral contaminate. It seems that when the live polio virus grown on monkey tissues was extracted for vaccine production another virus was extracted as well, SV-40. When this monkey virus was injected into research animals it produced brain cancer. It appears our government didn't wish to create a public panic or discredit the public health service, because instead of recalling the tainted vaccines, it quietly ordered the manufacturers to find a monkey free of SV-40 and continue production. As of 1963, the rhesus monkey had been replaced with the African green monkey for production of a safer polio vaccine, but between the years of 1955 and 1963 as many as 98 million Americans had received doses of live polio virus vaccines tainted with SV-40.
Jumping to the early 1990's, Michele Carbone, Assistant Professor of Pathology at Loyola University in Chicago, isolated fragments of the SV-40 virus in human bone cancers and in a particularly nasty form of lung cancer called mesotheliomas. The viral contaminate from the 50s was back to haunt us, and appeared in 33% of the osteosarcoma bone cancers studied, in 40% of other bone cancers, and in 60% of the mesotheliomas lung cancers. Dr. Carbone believed this study could explain why 50% of the current mesotheliomas being treated were no longer occurring in association with their traditional cause of asbestos exposure.
Already sounding like a bad science fiction story, the worse news was
yet to follow. An Italian team of researchers from the Institute of Histology
and General Embryology of the University of Ferrara lead by Dr. Fernanda
Martini discovered SV-40's presence in various other tumors.
To be specific they found the monkey virus in 83% of choriod plexus papillomas,
in 73% of ependymomas, in 47% of astrocytomas, in 50% of glioblastomas,
and in 14% of meningiomas.
While the virus's appearance in all of these types of brain tumors is
mortifying, even more so is the fact that it materialized in 23% of blood
samples and 45% of sperm fluids taken from normal individuals -- normal
meaning free of disease at the time of testing. The researchers determined
the virus could be transmitted sexually and through blood transfusions.
As if to drive this point home, SV-40 has appeared in 61% of all new cancer
patients -- patients too young to have received the contaminated vaccine
being administered forty years ago who are now believed to have been infected
by human to human transmission. Being a blood born organism, it is also
suspected that SV-40 is transmissible from mother to child during pregnancy.
The more this matter is researched the more startling the evidence. Senior epidemiologist at the National Institutes of Health, Dr. Howard Strickler, has plotted a geographic pattern to the cancers associated with SV-40 helping to confirm its link to the tainted vaccine. People who lived in Massachusetts and Illinois who received identified lot numbers of the contaminated vaccine administered in the 1950s are now demonstrating ten times the rate of the osteosarcoma bone tumors as those who received vaccine free of the SV-40 contaminate in other parts of the country.
The Food and Drug Administration (FDA) mandates that every American infant and child receive polio vaccinations. While public health officials continue to emphasize how current supplies of the vaccine are safe, Peter Reeve, FDA Virologist, has acknowledged that the administration abandoned independent testing of vaccine purity some fifteen years ago. The job of ensuring safety and purity rests squarely on the shoulders of those manufacturing the vaccines with no federal oversight. Wyeth-Lederle controls the supply of all the oral polio vaccine in this country, and last year's sales totaled some $230 million dollars. Surely there would be no conflict of interest in allowing this corporation to be the sole agent of quality oversight of their own pocketbook?
The government may not have paid attention to the quality of these vaccines, but they had formulated a plan for their distribution. Federal vaccination policy advocated the use of live-virus oral polio vaccine (OPV) based on the belief the live virus shed in the body fluids of infants immunized with OPV could immunize others through contact exposure. The Centers for Disease Control (CDC) insisted this was a safe practice, and emphasized that no one previously vaccinated could contract the disease in this manner. The public was never informed of this strategy, however, and no consent was ever obtained from the unknowing participants in this vaccination scheme. One hundred and twenty people, many previously vaccinated, contracted polio as a result of this practice. To add insult to injury in 1994 the World Health Organization proclaimed polio was eliminated from the Western Hemisphere. Insult because for the past seventeen years the only cases of polio occurring in the United States have been caused by the vaccine itself, and injury because this victory will be paid for in blood from the cancers produced by the monkey virus spread with the vaccine.
One might ask just how such a thing could happen considering the injectable form of the vaccine (IPV) does not use a live virus and doesn't transmit the disease it is designed to shield us from? Well, Wyeth-Lederle's leading competitor Connaught produces IVP which could explain why Wyeth lobbied so hard against the CDC recommending increased use of IVP. In 1996 the CDC revised its recommendation from four doses of OPV to two doses of IVP followed by two doses of OPV, however, physicians have been instructed to give all four doses as OPV if they desire. The cost of IVP vaccine is $5.40 per dose, whereas OPV costs $2.32 per dose. With the difference in cost favoring the use of OPV, and the current climate of regulating health care costs, clearer guidelines must come from the government if they truly expect to increase the use of the safer IVP vaccine.
Well the story of contaminated polio vaccine is not over yet. Microbiologist Howard Urnovitz, Ph.D. provided significant evidence at the Eighth Annual Houston Conference on AIDS that human immunodeficiency virus type 1 (HIV-1) is a monkey hybrid virus which was produced when 320,000 Africans were injected with polio virus contaminated with live simian immunodeficiency virus (SIV) in the late 1950's. Apparently, viral fragments combine easily with other viruses to produce these hybrids called "chimeras." This theory was confirmed by another research team headed by Dr. B. F. Elswood at the University of California in San Francisco. Interestingly enough, when researchers Cecil H. Fox and John Martin applied to the National Institutes of Health for grants to confirm the presence of SIV and simian cyto-megalovirus (SCMV) contaminates in polio vaccines their requests were denied. Dr. Urnovitz may have an explanation as he stated in the Boston Globe, "that almost 100 million Americans were exposed (to SV-40) through a government sponsored program, but for over 30 years, there has been virtually no government effort to see if anyone's been harmed by the exposure." He added, "The government will not fund science that makes it look culpable."
Could it be our government, once again, is attempting to avoid a public panic while ignoring the great potential for harm these viruses could inflict. Time will tell. Harvard Medical School professor, Dr. Ronald Desroier points out that taking all known scientific evidence into account that the medical experts' knowledge is limited to "perhaps 2% of existing monkey viruses." Who knows what lethal virus may be discovered in our blood streams forty years from now as a result of good intentions....
 |
Rife machines and Multiwave oscillators are claimed to complement each other based on the principle that life forms absorb energy. A multiwave Oscillator uses this principle to strengthen cells within the body to resist disease while a Rife machine uses this principle to destroy microorganisms with an overdose of frequency energy. |
 |
The worlds most advanved multi purpose Fully Automatic Rife/Hoyland System here |
 |
PEMF Pulser combo |
References
Berleur, M. P., & Cordier, S. (1995). The Role of Chemical, Physical,
or Viral Exposures and Health Factors in Neurocarcinogenesis: Implications
for Epidemiologic Studies of Brain Tumors.
Cancer Causes and Control, 6(3), 240-256.
Bookchin, D., & Schumaker, J. (1997). Tainted Polio Vaccine Still
Carries Its Threat 40 Years Later.
The Boston Globe, January 26.
Carbone, M., et al. (1996). SV-40 Like Sequences in Human Bone Tumors.
Oncogene, 13(3), 527-535.
Elswood, B. F., & Stricker, R. B. (1995). Polio Vaccines and the
Origin of AIDS.
Medical Hypotheses, 42(6), 347-354.
Fisher, B. L. (1997). Workshop on Simian Virus 40: A Possible Human Polyomavirus. National Vaccine Information Center, January 27, On-line at http://www.909shot.com/polio197.htm>http://www.909shot.com/polio197.htm.
Krieg, P., Amtmann E, Jonas, D., Fischer, H., Zang, K., & Sauer G.
(1981). Episomal Simian Virus 40 Genomes in Human Brain Tumors.
Proceedings of the National Academy of Sciences of the United States of
America, 78(10), 6446-6450.
Lednicky, J. A., Garcea, R. L., Bergsagel, D. J., & Butel, J. S.
(1995). Natural Simian Virus 40 Strains are Present in Human choroid Plexus
and Ependymoma tumors.
Virology, 212(2), 710-717.
Martini, F., et al. (1995). Human Brain Tumors and Simian Virus 40.
Journal of the National Cancer Institute, 87(17), 1331.
Martini, F., et al. (1996). SV-40 Early Region and Large T Antigen in
Human Brain Tumors, Peripheral Blood Cells, and Sperm Fluids From Healthy
Individuals.
Cancer Research, 56(20), 4820-4825.
Pass, H. I., Kennedy, R. C., & Carbone, M. (1996). Evidence for and
Implications of SV-40 Like Sequences in Human Mesotheliomas.
Important Advances in Oncology, 89-108.
Rock, A. (1996). The Lethal Dangers of the Billion Dollar Vaccine Business.
Money, December, pages 148-163.
Tognon, M., et al. (1996). Large T Antigen Coding Sequences of Two DNA
Tumor Viruses, BK and SV-40, and Nonrandom Chromosome Changes in Two Glioblastoma
Cell Lines.
Cancer Genetics and Cytogenics, 90(1), 17-23